This post summarizes work conducted at both Wild Animal Initiative and Animal Ethics. The first part can be found here and continues here.
Introduction
The documentary series "Our Planet" opens with a flamingo chick whose legs have become caked with salt from the mud flats. The young bird can't keep up with the rest of their flock and is left to die. At the same time, other chicks and healthy adults seem to be living reasonably contented lives, able to find food and overcome other challenges. It seems plausible that an adult flamingo has lived a life characterized more by pleasure than suffering. This chick, though – and some proportion of all flamingos who have been born – never got to experience their best years.
As part of an ongoing project to understand the welfare of wild animals, I analyzed age-specific mortality rates and considered how they might relate to welfare, introducing a new concept for understanding the lives of wild animals: welfare expectancy. Welfare expectancy can serve as a framework for weighing up the different levels of well-being animals might experience over the course of their lives, helping to model the welfare consequences of interventions and natural pressures, such as predation, that may disproportionately affect animals of particular ages. The pre-print can be found here, and the paper is currently under review.
Welfare expectancy
To understand the balance of pleasure and suffering in nature, we need to understand what proportion of animals experience different welfare-relevant outcomes. In the face of the diversity of individual experiences wild animals may have, we need a way to tally them up in order to assess the overall welfare of a population. Expected value does this by taking the sum of the value of each outcome multiplied by its probability. For example, to calculate the life expectancy of a population (i.e. the expected value of lifespan), one would multiply the proportion of individuals who die at a certain age by the number of years they lived and sum this across all possible lifespans.
Lifespan is a blunt way of quantifying welfare outcomes, because two animals may have very different experiences and yet die at the same age. However, within the same species, the frequency of different lifespans is likely to reflect common challenges associated with specific stages of life. Since only living animals are capable of experiencing welfare, lifespan is effectively an upper bound on the amount of affectively positive or negative experience an animal can accrue.
If average welfare levels are constant throughout life, then life expectancy is the only welfare-relevant metric we can derive from patterns of age-specific mortality. However, welfare likely does vary with age, as juveniles, sub-adults, reproductive adults and senescent animals face different levels and forms of disease, competition, predation and environmental hardship. This potential for variation calls for a distinct concept of welfare expectancy.
Consider a species for which welfare is poor in early life, but high in adulthood. If the probability of surviving early life is high, then the lifetime expected value of welfare for an individual born into that population may be high, because most individuals are going to have a chance to live out their best years in adulthood. If the probability of surviving early life is low, then most individuals will only live to experience the juvenile period of poor welfare. Conversely, in a species where welfare is higher in early life than in adulthood (e.g. due to good parental care), the net welfare of even a short-lived animal could be relatively high.
We are profoundly uncertain about whether most animals' lives are dominated by pleasure or suffering, or even how to go about weighing these up. Therefore, it may be prudent to concentrate on a measure of "relative welfare expectancy" (RWE), representing the normalized welfare expectancy of a population divided by its life expectancy. For a fixed life expectancy, the highest welfare expectancy is achieved by maximizing the proportion of animals living to experience the best years of life while minimizing the proportion experiencing the worst years.
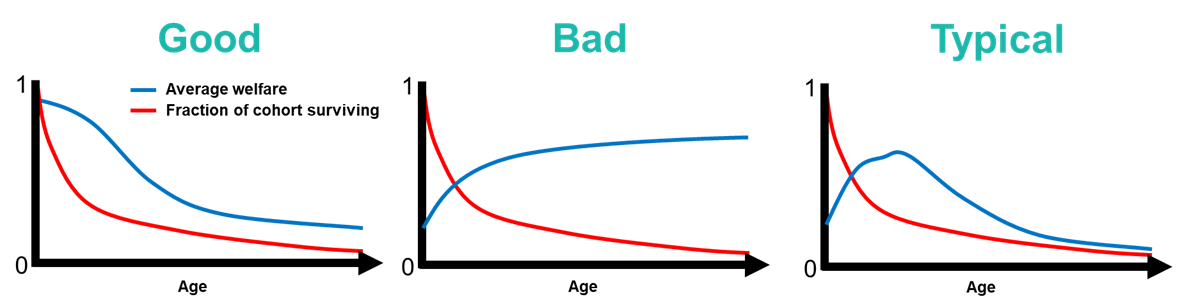
Confident application of the welfare expectancy concept will require empirical data on values of age-specific welfare, which arecurrently scarce for wild animal populations. A plausible working hypothesis, however, is that the average welfare experienced by an animal of a given age is proportional to their probability of surviving that period of life. The justification for this is that the same factors which lead to mortality (e.g. disease, vulnerability to predators, competition for food) have been shown to lead to chronic stress and poor physical condition.
Age-specific mortality
The vast majority of animals live very short lives; not only in absolute terms, but also relative to the longest-lived members of their species. In fact, of the populations analyzed in the paper associated with this project, average lifespans were on average 16% of a species' maximum lifespan, with only 5% of populations having life expectancies >33% of their maximum. Importantly, this represents an average across populations, not across individuals. Because short-lived species tend to be more populous, lives in nature are likely to be cut short far more often than these numbers suggest. Depending on how welfare varies with age in their respective species, especially short-lived individuals will be missing out on a great deal of positive and/or negative experience.
Patterns of age-specific mortality are too diverse to assign universal classifications to large taxonomic groups. For example, even some insect species have relatively high rates of juvenile survivorship. However, while age-specific patterns vary, animals of certain groups, such as the ray-finned fishes (actinopterygii), do have considerably shorter life expectancies and lower average annual survival rates than others, including birds (aves) and mammals. Importantly, life expectancy is only an average of the lifespans. As explained above, not only the average, but the precise distribution of lifespans is important to consider because of how it may correspond to the distribution of age-specific welfare. For a detailed discussion of some examples of age-specific mortality patterns, see the post by Animal Ethics or the pre-print manuscript.
For an animal to have an enjoyable life on net, they must experience enough pleasure to compensate for the pain of their death. Cause of death, and therefore the duration and pain of an animal’s experience of dying, may also vary with age similarly to welfare. In a hypothetical species, juveniles might be most likely to starve while adults are most likely to be predated, with the relative probabilities of these and other mortality factors shifting over a lifetime. If the pain of death is a sufficiently strong factor to negate some of the positive welfare an animal might have experienced while alive, age-specific variation in the incidence of various manners of death and their severity could also be important to account for.
Conclusion
At the individual level, welfare expectancy unites two distinct concepts: day-to-day quality of life and the quantity of welfare experienced over an individual’s lifetime. However, a similar quantity-quality distinction applies at the level of populations, with welfare expectancy addressing the quality side of the argument and quantity being determined by the population size. Ideally, a population should be managed in such a way that maximizes its total welfare expectancy.
The field of welfare biology is at a very early stage, and little dedicated work from the life sciences has been invested up until recently. While progress is still limited by the lack of empirical studies of wild animal welfare, knowledge of age-specific mortality patterns and a predictive understanding of population ecology will be essential for contextualizing this information, as well as evaluating and prioritizing among interventions that differentially affect various age groups.


I found this comment interesting "Ideally, a population should be managed in such a way that maximizes its total welfare expectancy." and wonder how you envision that proceeding? For example, for some large groups of herbivorous insects, the juvenile life stage is probably the best in terms of welfare Cuddington 2019 (and in some cases, the longest stage), but an optimal strategy might therefore imply that we should prevent these individuals from maturing and experiencing the harms associated with death by exposure etc etc. But of course, result of that strategy would be that a given species would become extinct in about one generation. Related to this, I also wonder how one reconciles managing a given population for optimal welfare given the downstream welfare effects on other species (mutualists, competitors, predators, amenalists, keystone species etc etc).
I completely agree that the downstream effects on other species should be accounted for. The true ideal would be to manage a population in such a way that maximizes the total welfare expectancy of all sentient life! I hope we'll eventually have the information necessary to do so. In the near term, I'm advocating for populations that are currently managed in such a way as to maximize their net reproductive rate (i.e. 'fecundity expectancy') to instead be managed around total welfare expectancy.
Given the relevant information, I think the welfare expectancy approach makes sense of the example of an herbivorous insect where the juvenile stage offers the highest welfare, but where locking them into that stage would lead to rapid extinction.
(edited to correct numbers:) For example, assume that the average survival rate and welfare during the juvenile stage is 0.9 and during the adult stage is 0.2; the juvenile stage lasts ~1.8 months while adults survive a further ~0.7 months; and adults produce an average of 5 offspring per month. Assume we could determine a proportion of the larvae to develop into adults while the rest remain juveniles, and that this wouldn't affect any survival/welfare/fecundity rates. The lifetime welfare expectancy of juvenile-locked individuals would be ~8.08 and of "normal" individuals ~1.56. "Normal" individuals would produce an average of 4.86 offspring over their lifetime. A higher proportion of individuals being limited to the juvenile stage increases average welfare expectancy per individual, but curtails the population size. A lower proportion of individuals being limited to the juvenile stage means average welfare expectancy is lower, but the number of individuals experiencing it is higher.
Plugging in the numbers above, I find the total welfare expectancy is maximized when 38% of individuals are limited to the higher-welfare (but non-reproductive) juvenile stage. Of course, the assumption that such an intervention wouldn't affect age-specific survival/welfare rates seems implausible, and this still isn't accounting for potential effects on other species. However, I hope this illustrates how the usefulness of this approach to wild animal welfare might scale with better ecological understanding!
I totally get where you are going with this.... but do want to offer the following observation regarding the context dependence of life-stage associated welfare. Many of these species have tight seasonal links for development that in turn are related to factors like plant phenology. So.... just to make it more specific, for the things like juvenile leaf miners (ample food, fairly but not completed protected from predators and parasitoids,) you might "lock" individuals into a declines in both food quality and quantity, and increases in predation and parasitism by preventing them from emerging from the leaf as the the season advances. Thus decreasing the welfare of those "locked" individuals.