I absolutely love this. The data visualisation is beautiful, the collection of cognitive and hedonic proxies is comprehensive and there is a source link for every cell where the response isn't "unknown".
I've just checked your welfare range table and it's a pretty amazing work you did there and the illustration is gorgeous :)
I'm a French science populariser and I'm mostly specialised into farm animals and fishes. Here are a few thoughts I had when reading here and there, especially the Excel file, if ever they could be of any use to you :)
Maybe adding sheep might be a good idea as most of the studies on the appraisal theories framework have been done on them and that could add a few things. Veissier and Boissy's work and Lucile Greiveldinger thesis have very interesting things.
The last part of her thesis has a non-published study (but I have it, if you need it) of the influence of the social rank in the expression of the emotions in sheep and actually I'm quite sure I have that also in at least one fish specie, if ever you could be interested.
Sheep is also the specie with one of the most impressive known self-medication abilities we have today.
On hen we also have the "higher" level of empathy, cognitive empathy. In your references is listed the "lower level one", it might be worth adding this paper which is much more exceptional : https://doi.org/10.1016/j.anbehav.2013.05.004
You might also want to add trout or make a salmonid (as you already have an "octopus" or "bee" case for several species) because we have studies on trace conditioning as well there https://link.springer.com/article/10.1007/s10071-009-0267-3 (as far as I know, we have shows trace conditioning in a few other species like cod, and as salmon and trout are pretty close and we have a few other very interesting stuff on trouts... ^^)
I've noticed in the references table that you often have only one paper, I don't know if it's done on purpose but there are often much more to backup your position (for example in bee judgement bias, there's a pessimist but also an optimist bias paper https://www.science.org/doi/10.1126/science.aaf4454 )
I'm looking forward to see where all this goes ! Far, I hope! Nice job :)
This is great feedback, Sebastien! Thanks so much for all the recommendations. We do want to keep updating the table with new references and, eventually, expand it to include other species. Cattle, sheep, and goats are probably the most important terrestrial vertebrates, but many aquatic animals and invertebrates are farmed in much greater numbers. Hopefully, we'll be able to include them all over time.
As for the decision to include just one paper, that was mostly to keep the table relatively neat. Moreover, if one paper makes a decent case for the presence or absence of a trait, it isn't clear whether our credences would change enough, given the large "credence buckets" we're using, to justify including more of the literature. But I agree that, all else equal, it would be better to have as many references as possible.
Nice! (admit I've only just skimmed and looked at the eye-catching graphics and tables 🙃). A couple small potential improvements to those things:
Is a higher-quality/bigger file version of the infographic available? Shouldn't matter, of course, but may as well put it on a fair memetic playing field with all the other beautiful charts out there
Would you consider adding a few "reference" columns to the Welfare Range Table, in particular values for:
Human (or perhaps "human if introspection is epistemically meaningful and qualia exist")
Organisms from other kingdoms: plant, bacteria, etc. (I think any single one without nervous tissue would suffice)
A representative (very probable) non-moral patient physical object ("rock")
Less important (intuitively to me) potential additions:
chatGPT
Video game character or anything else that "behaves" in some sense like an advanced organism but is ontologically very different
any other contrarian counterexample things that might push the limits of the taxonomy's applicability (maybe 'Roomba' or 'the IOS operating system' or 'a p-zombie' ?)
I never curated this when it first appeared, but I'm curating it now to give people an extra nudge to read or skim the post and explore the methodology. And of course, the visualizations are excellent. (Thanks so much for working on this project, all!)
One reason I'm curating this right now is that the discussion around Why Neuron Counts Shouldn't Be Used as Proxies for Moral Weight (both in the comments and in conversations I've had about the post since) seems to often center on considerations like, "ok, neuron counts are great, but we need to use something and it's not clear what to use." I think this post offers a concrete counter-proposal.
Assorted other things I like (non-exhaustive list):
Being clear on the limitations of the information presented (there are very few negative results, etc.), or potential biases that might have influenced it
The spreadsheet is really well presented, and I really appreciate the fact that you've given readers the chance to explore it and follow the sources
Maybe this is obvious, but the core subject of the post is extremely important and a point of uncertainty or disagreement with a lot of people. That's great.
Thank you, Lizka! We really appreciate your drawing attention to our work. In January, we’ll propose a way to aggregate all this information to generate welfare range estimates. In the interim, there’s a bit more ground-clearing to do.
We don't, I'm sorry to say. The numbers would be comparable to pigs, but because cows are farmed in such low numbers by comparison, we didn't prioritize them. I know we need to extend the analysis, given how many people have asked about cattle!
Got it, thanks. I'm interested in the cattle analysis because cows yield ~4x more meat than pigs per slaughter, and could perform even better than that when factoring in cognition.
Really interesting. I've acted as though insects were sentient out of moral caution, and I'm surprised to see there is more research and more confidence on them than I thought. Thanks for this compilation!
Thank you so much for putting this together, I really appreciate seeing this type of work getting such serious attention! Sorry if I'm missing something obvious, but are the sources compiled somewhere? I thought that maybe they would be linked in the spreadsheet, but at least for me whenever I click on a hyperlink in the spreadsheet it just opens the same sheet in a different tab. Thanks!
Sorry about that! I've fixed the link, which now goes directly to the Google Sheet. Originally, you would have had to scroll down to switch tabs to see the references. Now, clicking through will work. Thanks for bringing this to my attention!
I think it would be great if you added nematodes to the graphs. You ended up estimating their probability of sentience to be 6.8 %, 82.9 % (= 0.068/0.082) of the 8.2 % of silkworms, which you included in the graphs.
Did you end up estimating a probability of sentience for plants, which you also included in your sentience table? It would be interesting to know how it compares with that of nematodes.
Did you end up estimating a probability of sentience for plants, which you also included in your sentience table? It would be interesting to know how it compares with that of nematodes.
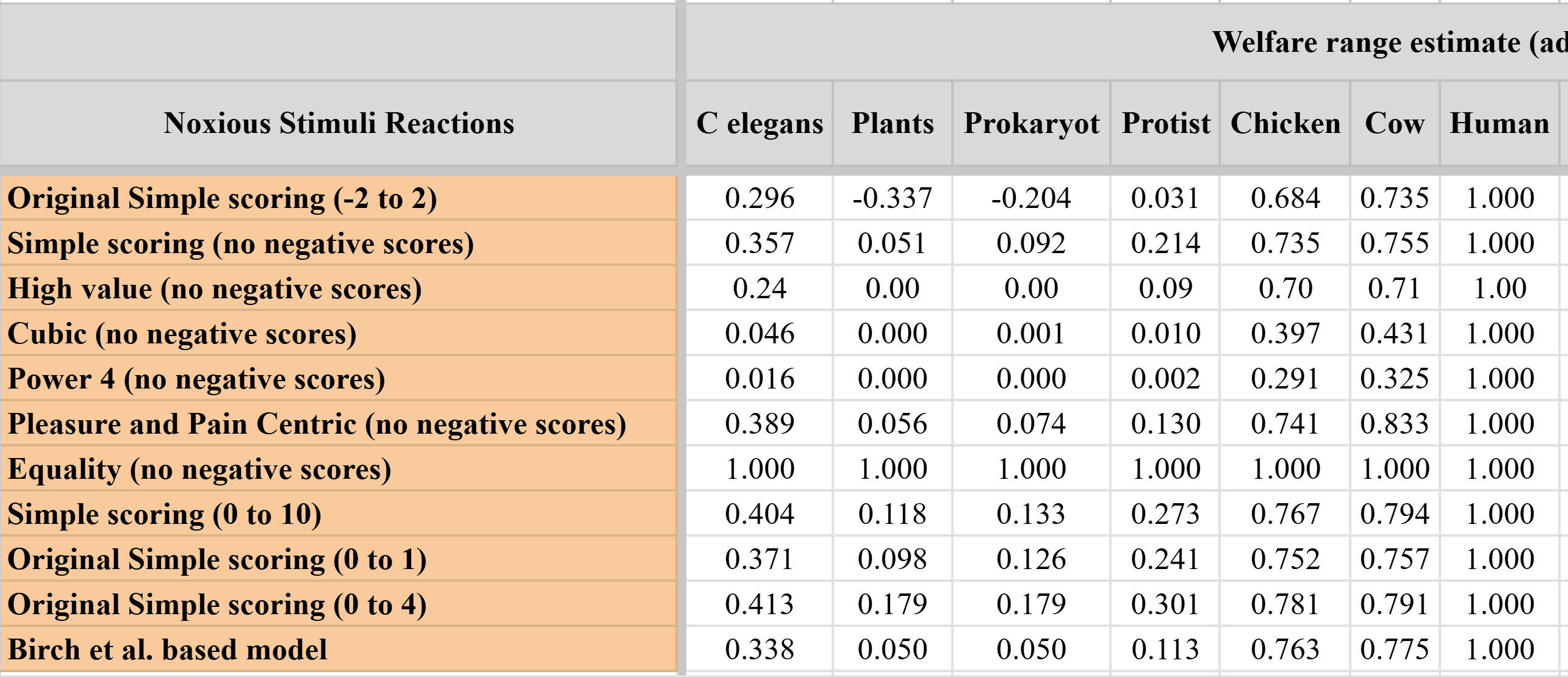
I do not remember where I found the sheet I linked above, but the 1st tab has the estimates below for plants, nematodes, and microorganisms supposedly referring to "Welfare range estimate (adjusted for flicker fusion frequency)" which look super high. Do the estimates really refer to welfare ranges? If so, are they supposed to account for the probability of sentience, or flicker fusion frequency[1]? The cubic, and pleasure and pain centric models were 2 of the 9 models you used to estimate your mainline welfare ranges. If the values below refer to welfare ranges accounting for the probability of sentience, I am confident that you would get mainline welfare ranges of microorganisms sufficiently high for effects on them to dominate all else.
If the values below refer to welfare ranges accounting for the probability of sentience, I am confident that you would get mainline welfare ranges of microorganisms sufficiently high for effects on them to dominate all else.
This post presents the executive summary from Giving What We Can’s impact evaluation for 2025. At the end of this post we share links to more information, including the full report and...
I used AI to fix transcription errors, rerrarange the ideas, and suggest tweaks to the title and some sentences.
Three of the most exciting projects to come out of EA in recent years are, in a vague sense, CEA spinouts:
* Kairos is directly a spinout of CEA and now handles most support for university AI safety groups. Basically everyone I've found who knows them is really excited about what they do
* NEST is an opinionated ideas-fi...
Our objective: estimate the welfare ranges of 11 farmed species.
Given hedonism, an individual’s welfare range is the difference between the welfare level associated with the most intense positively valenced state that the individual can realize and the welfare level associated with the most intense negatively valenced state that the individual can realize.
Given some prominent theories about the functions of valenced states, we identified over 90 empirical proxies that might provide evidence of variation in the potential intensities of those states.
There are many unknowns across many species.
It’s rare to have evidence that animals lack a given trait.
We know less about the presence or absence of traits as we move from terrestrial vertebrates to most invertebrates.
Many of the traits about which we know the least are affective traits.
We do have information about some significant traits for many animals.
Introduction
This is the second post in the Moral Weight Project Sequence. The aim of the sequence is to provide an overview of the research that Rethink Priorities conducted between May 2021 and October 2022 on interspecific cause prioritization—i.e., making resource allocation decisions across species. The aim of this post is to provide an overview of the Welfare Range Table, which records the results of a literature review covering over 90 empirical traits across 11 farmed species.
Motivations
If we want to do as much good as possible, we have to compare all the ways of doing good—including ways that involve helping members of different species. The Moral Weight Project’s assumptions entail that everyone’s welfare counts the same and that all welfare improvements count equally. Still, some may be able to realize more welfare than others. We’re particularly interested in how much welfare different individuals can realize at a time—that is, their respective welfare ranges. An individual’s welfare range is the difference between the best and worst welfare states the individual can realize at a time. We assume hedonism, according to which all and only positively valenced states increase welfare and all and only negatively valenced states decrease welfare. Given as much, an individual’s welfare range is the difference between the welfare level associated with the most intense positively valenced state that the individual can realize and the welfare level associated with the most intense negatively valenced state that the individual can realize. In the case of pigs, for instance, that might be the difference between the welfare level we associate with being fully healthy on a farm sanctuary, on the one hand, and a botched slaughter, on the other.
If there’s variation in welfare ranges across taxa, then there’s variation in the capacities that generate the determinants of welfare. So, if there’s such variation and hedonism is true, then there’s variation in the capacities that generate positively and negatively valenced experiences.
As Jason Schukraft argues, we don’t have any good direct measures of the intensity of valenced states that let us make interspecific comparisons. Indeed, we rely on indirect measures even in humans: behavior, physiological changes, and verbal reports. We can observe behavior and physiological changes in nonhumans, but most of them aren’t verbal. So, we have to rely on other indirect proxies, piecing together an understanding from animals’ cognitive and affective traits or capabilities. The Welfare Range Table includes over 90 such traits: some behavioral, others physiological; some more cognitive, others more affective. Then, it indicates whether the empirical literature provides reason to think that members of 11 farmed species possess those traits.
Methodology
Our team was composed of three philosophers, two comparative psychologists (one with expertise in birds; another with expertise in cephalopods), two fish welfare researchers, two entomologists, an animal welfare scientist, and a veterinarian. To select proxies for variation in the capacities that generate positively and negatively valenced experiences, we needed to rely on general theoretical considerations. In this case, we relied on theories about the function of valenced experiences—that is, their adaptive value for organisms who have them.
There are three main theories about the function of valenced experiences. The first is that they allow organisms to represent fitness-relevant information (“That’s good” / “That’s really good” / “That’s bad” / “That’s really bad” / etc.; Cutter & Tye 2011). The second is that they provide a common currency for decision-making (“A is better than B”; Ginsburg & Jablonka 2019). The third is that they facilitate learning (“If X, then A” / “If Y, then B”; Damasio & Carvalho 2013). So, we looked for empirical traits that arguably bear some relation to information representation, decision-making, and learning via valenced experiences. You can find an annotated list of those traits here.
For each taxon / trait pair, we began with searches via Google Scholar. Then, we looked at the references in recent reviews. If none of those sources provides positive or negative evidence, we searched via Web of Science. Then, we broadened the search. If, for instance, the target trait hadn’t been studied in the relevant species, we looked at the family; if the trait hadn’t been studied in the relevant family, we looked at the order. If it seemed plausible that the results of the study generalize to the target species—for example, because of other shared traits or similar ecological niches—then we included those results as well.
Based on the evidence available for each taxon and trait, we made one of five assessments:
Likely No (Credence: 0% – 25%)
Lean No (Credence: >25% – <50%)
Unknown (Credence: 50%)
Lean Yes (Credence: >50% – 75%)
Likely Yes (Credence: >75% – 100%)
These broad credence ranges are about trait possession (do members of the taxon have the trait or not?), not degree of possession (relative to humans, to what degree do the members of this taxon possess this trait?). The assessments are based on the existing published scholarly literature (as of August 2022), with “Unknown” being the default assessment in cases where there was no literature that spoke fairly directly to a given taxon / trait pair.
After completing the literature reviews, we performed two quality checks. First, we checked for consistency. In our initial review, for instance, honey bees got a “lean yes” for transitive inference, whereas pigs got a “lean no.” A closer look revealed that researchers were holding these animals to different standards. (In this particular case, we opted to flip pigs to “lean yes,” as the lower standard seemed reasonable for our purposes.) Second, we commissioned spot checks of 15% of the traits per taxon from someone who had not been involved with the initial literature review for that taxon. Across all the taxon / trait combinations, the spot checkers didn’t flip any positive responses (“likely yes”; “lean yes”) to negative responses (“likely no”; “lean no”) or vice versa. However, they did flip a handful of “unknown” responses to “lean yes.”
Results
The Welfare Range Table is available here. However, the following diagrams provide a more digestible visual representation of most of the information we gathered (excluding physiological proxies):
We should highlight a few key results. First, as these diagrams suggest, there are many unknowns. This shouldn’t be surprising: in many cases, no one has been interested in investigating these traits; in many other cases, no one knows how to investigate them. Moreover, insofar as it’s easier to publish positive results than negative results, academics have an incentive to study traits that they already expect to find. So, the literature is probably biased toward relatively “sure things,” in the interest of guaranteeing that people avoid walking away from their research empty-handed (i.e., without publications). This point is relevant to—and partially explains—the second key result, which is that there are very few negative results.
The table below indicates the percentage of proxies for which we were able to find some information, whether positive or negative. As the table indicates, information of any kind generally declines from terrestrial vertebrates to invertebrates.
As the next table makes clear, many of the traits about which we know the least are affective.
On one level, this shouldn’t be surprising either, given scientists’ determination not to anthropomorphize animals—i.e., attribute human characteristics to nonhumans. (Unfortunately, scientists seem to worry less about what Franz de Waal calls “anthropodenial”—i.e., “the a priori rejection of shared characteristics between humans and animals.”) Still, these results are concerning insofar as we think that the welfare impacts of valenced states are additive, where the welfare impacts of experiences “stack” on top of one another. On such a view, experiencing maximal pain and shame (for instance) is worse than experiencing maximal pain alone. So, if we don’t know whether animals can experience shame, we don’t know something that bears directly on their welfare ranges.
However, it might not be the case that valenced states are additive: it could be that when you’re experiencing maximal pain, you’re unable to experience anything else, as pain consumes all your attention. If that’s right, then while information about many valenced states might be highly relevant to assessing welfare impacts (how animals are actually faring), it isn’t necessarily important for assessing welfare ranges (how animals could fare), at least if pain states are the most intense negatively valenced states that animals can experience.
That said, it isn’t completely obvious that pain states are the most intense negatively valenced states that animals can experience. So, that provides some motivation for attending to other kinds of negatively valenced states. Moreover, while we’re skeptical that negative experiences are additive, we aren’t certain, so we assign some credence to the additive view. In expectation, then, we take information about the breadth of animals’ emotional lives to be relevant to their welfare ranges.
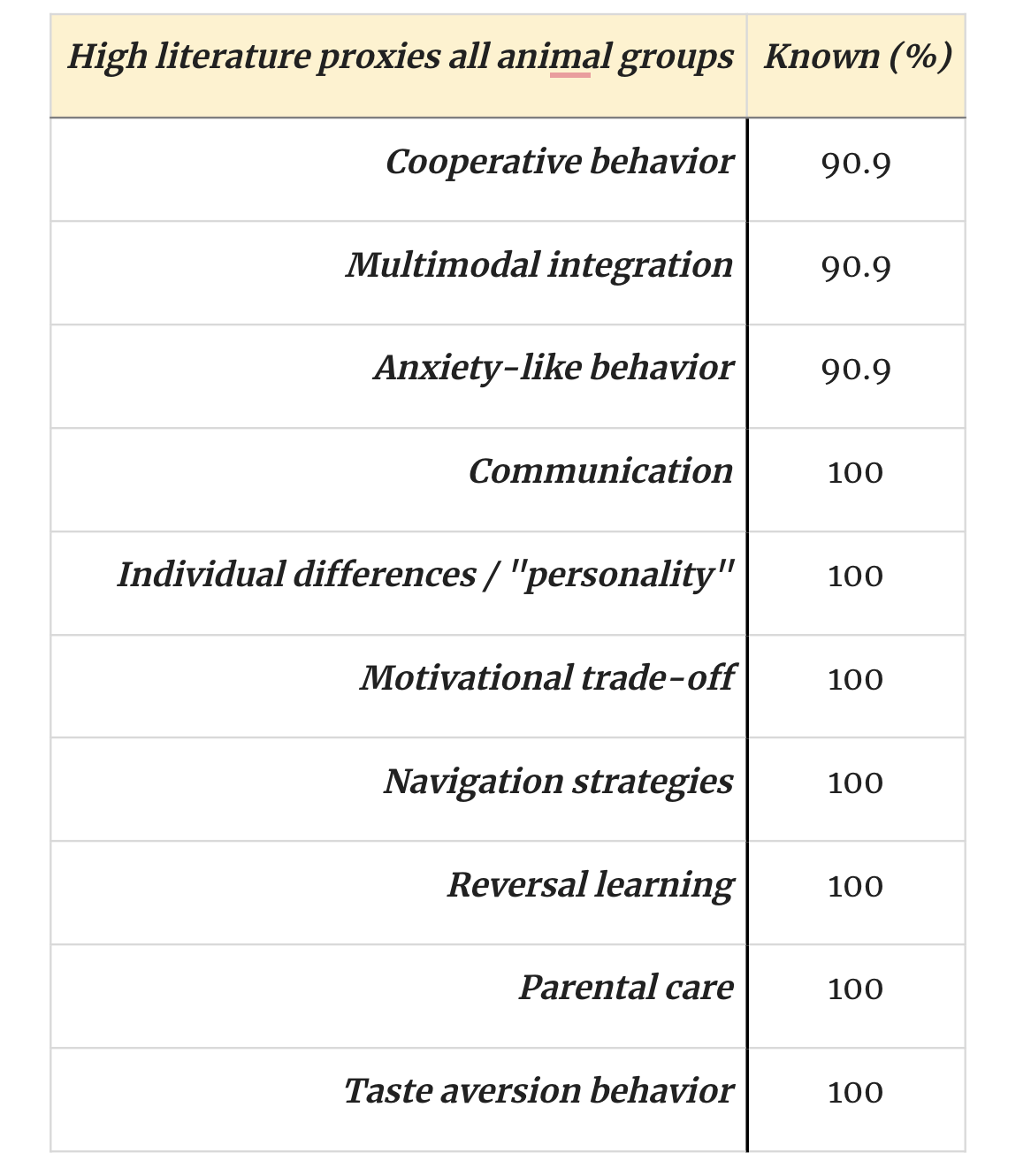
In any case, one final point to note is that we do have information about some significant traits for many animals, as the following table indicates:
Parental care, for instance, provides some evidence of the range of affective states available to an organism, as does anxiety-like behavior. Multimodal integration is often touted as a key piece of evidence for sentience, as is the capacity to make trade-offs. But both traits are also relevant to welfare ranges insofar as they come in degrees (as suggested by the presence or absence of other traits).
A clarification and some limitations
To be clear: the Welfare Range Table does not automatically supply welfare range estimates. It’s one thing to collect data that are relevant to welfare ranges; it’s another thing to score and aggregate them. In this post, we’re only discussing the data. We’ll defer the discussion of scoring and aggregation until later in the Moral Weight Project Sequence.
The Welfare Range Table is the first of its kind. Inevitably, then, it has several limits. Among them:
Our searches in related taxa were limited. In the interest of completing the project in a reasonable timeframe, we tried to limit the search for each taxon / trait pair to one hour. In fact, we often spent two or three hours per taxon / trait pair. Still, spending more time would have made it possible to investigate a wider range of related taxa for each taxon / trait pair.
The proxy list could be expanded. We opted for an inclusive approach to the proxies. This made the project enormous. Still, there are many other traits that could have been included—and, in some cases, perhaps ought to have been included in a list of this length.
The proxy list could be refined. There are two senses in which the proxy list could be refined. First, some proxies may not be sufficiently informative about animals’ welfare ranges—though, of course, this is hardly obvious, as we don’t have an independent way to assess how informative they are. Second, many of the proxies are relatively coarse-grained. Consider a trait like reversal learning: namely, the ability to suppress a reward-related response, which involves stopping one behavior and switching to another. This trait comes in degrees: some animals can learn to suppress a reward-related response in fewer trials; and, having learned to suppress a reward-related response at all, some can suppress their response more quickly. A more sophisticated version of the table would account for this variation.
Academics are biased toward “Unknown.” We thought it was important to have domain experts review the literature whenever possible. However, domain experts are academics. Academics are socialized into a community where it’s inappropriate to make some positive claim (“Pigs have this trait” or “pigs lack that trait”) without being able to establish that claim to the satisfaction of your peers. There are good reasons to value this socialization in the present case. For instance, it’s difficult to predict which traits an organism will have based on its other traits. Moreover, it’s difficult to predict whether one kind of organism will have a trait because a related kind of organism does. Still, even though the probability ranges we mentioned earlier establish a very low bar for “lean yes” and “lean no” (above and below 50%, respectively), we defaulted to “unknown” when we couldn’t find any relevant literature. Even if our approach is defensible, other reasonable literature reviewers may have had more “lean yes” and “lean no” assessments than we did.
We could address these issues in the future. With enough time, for example, we could make inferences about trait possession from more distant taxa. Or, we could make efforts to include any neglected high-value proxies, eliminate relatively low-value proxies, and refine the proxies insofar as that’s possible. Finally, we could try using a missing data method to account for systematic trends in the “Unknowns.”
Conclusion
The Welfare Range Table is the first attempt to assess a wide range of welfare-relevant traits in such a diverse array of taxa. It represents an initial but significant step toward estimating the differences in animals’ welfare ranges. If it proves to be valuable, then it ought to be extended, as we didn’t examine many important species. While we prioritized many of the most-farmed taxa, there are many others that humans farm in extraordinary numbers. In particular, it would be good to investigate more fishes and invertebrates.
Acknowledgments
This research is a project of Rethink Priorities. It was written by Bob Fischer. Thanks to Jason Schukraft and Adam Shriver for much of the conceptual work behind the project. Thanks to Rachel Norman, Martina Schiestl, Alex Schnell, and Anna Trevarthen for helpful feedback on earlier versions of this post. Thanks to Meghan Barrett, Leigh Gaffney, Michelle Lavery, Rachael Miller, Martina Schiestl, Alex Schnell, and Anna Trevarthen for their extensive literature reviews. Thanks to Jamie Elsey for the visuals. If you’re interested in RP’s work, you can learn more by visiting our research database. For regular updates, please considersubscribing to our newsletter.



 We should highlight a few key results. First, as these diagrams suggest, there are many unknowns. This shouldn’t be surprising: in many cases, no one has been interested in investigating these traits; in many other cases, no one knows how to investigate them. Moreover, insofar as it’s easier to publish positive results than negative results, academics have an incentive to study traits that they already expect to find. So, the literature is probably biased toward relatively “sure things,” in the interest of guaranteeing that people avoid walking away from their research empty-handed (i.e., without publications). This point is relevant to—and partially explains—the second key result, which is that there are very few negative results.
We should highlight a few key results. First, as these diagrams suggest, there are many unknowns. This shouldn’t be surprising: in many cases, no one has been interested in investigating these traits; in many other cases, no one knows how to investigate them. Moreover, insofar as it’s easier to publish positive results than negative results, academics have an incentive to study traits that they already expect to find. So, the literature is probably biased toward relatively “sure things,” in the interest of guaranteeing that people avoid walking away from their research empty-handed (i.e., without publications). This point is relevant to—and partially explains—the second key result, which is that there are very few negative results.


I absolutely love this. The data visualisation is beautiful, the collection of cognitive and hedonic proxies is comprehensive and there is a source link for every cell where the response isn't "unknown".
Thanks for the kind words, Emre!